天津师范大学学报·自然科学版杂志投稿格式参考范文:嗜水气单胞菌非编码RNA的功能研究
时间:
1 材料与方法
1.1 实验材料
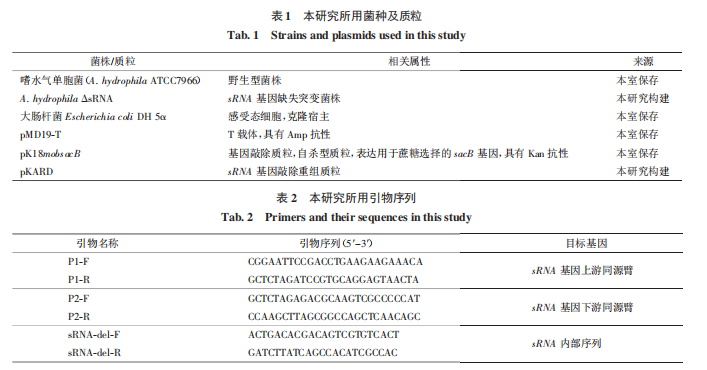
本研究所用菌种及质粒见表 1。LB(Luria-Bertani)培养基用于大肠杆菌和嗜水气单胞菌的培养,购自赛默飞世尔科技公司;绵羊血琼脂平板购自北京索莱宝科技有限公司;限制性内切酶(EcoR I、Xba I 和 Hind Ⅲ)、T4 DNA 连接酶、DNA Marker、FastPure Plasmid Mini Kit 质粒小量提取试剂盒,均购自 Takara 公司;基因组提取试剂盒,购自全式金有限公司;琼脂糖,购自 Promega 公司;Agar Powder,购自 Solarbio 公司;Gel Extraction Kit,购自 OMEGA 公司。氨苄青霉素(Amp)和卡那霉素(Kan)贮存液终质量浓度为 100 mg/mL。BALB/c 小鼠购自河北医科大学动物中心。本研究所用引物由金唯智生物技术公司合成。
1.2 实验方法
将嗜水气单胞菌接种于 LB 液体培养基中,30 ℃、180 r/min 过夜培养;4 500 r/min 离心 10 min,弃上清,收集菌体。根据 TRIzol® 试剂盒说明书提取 A.hydrophila 的总 RNA,使用 Nanodrop1000 微量紫外分光光度计(赛默飞世尔科技公司)和 1% 琼脂糖凝胶电泳测定 RNA 的质量浓度(ng/μL)、纯度((OD260/OD28))和片段大小,利用 Agilent 5400 片段分析仪(Agilent Technologies 有限公司)测定 RNA 完整性值(RIN)。确定 RNA 浓度是否满足建库测序质量要求。
RNA 文库构建、质控和测序:原核生物转录组测序采用去除 rRNA 链特异性的建库方式,严格要求测序数据质量(Q20>90%,Q30>85%);库检合格后,用 NovaSeq 6000 测序平台(美国 Illumina 公司)进行高通量测序,测序读长为 PE150。
转录组测序数据分析:首先对测序数据质量进行评估,过滤低质量数据,保证数据质量,与嗜水气单胞菌参考基因组进行比对,分析对比率;其次对样本的基因表达水平进行分析来评估该样品中基因的表达量;最后基于以上标准分析,对该样品进行原核转录组的高级分析。分析主要包括 SNP/Indel 分析变异位点、挖掘新的基因或转录本、对操纵子、转录起始位点和终止位点等基因结构分析以及对 5′UTR SD 序列预测和 3′UTR 终止子序列预测,此外分析该样品反义转录本在基因组上的位置、种类以及数量等。将经过组装的测序结果与已注释的基因模型进行比较,应用 Rockhopper 软件发现新的基因区间转录本,通过 Blastx 与 nr 库作比对,对新预测的转录本区域进行注释,将注释不上的转录本作为候选的非编码 sRNA。
对预测出的 sRNA 进行二级结构预测和靶基因预测。用 TargetRNA 2.01 软件(https://cs.wellesley.edu/~btjaden/TargetRNA2/index.html)对候选的 sRNA 的靶基因进行预测,以 Aeromonas hydrophila subsp. hydrophila ATCC 7966 chromosome 作为数据库。利用 BLAST 挖掘其他物种中的 sRNA,然后使用 ClustalX 软件和 GenDoc 软件进行多序列比对,分析 sRNA 在不同物种中的保守性。
37 ℃、180 r/min 过夜培养复苏菌液后提取 A.hydrophila 基因组。以基因组 DNA 为模板,分别用 P1FP1-R 和 P2-FP2-R 引物进行 PCR,扩增 sRNA 上游同源臂(660 bp)和下游同源臂(880 bp)。反应程序:95 ℃ 3 min,95 ℃ 15 s,55 ℃ 30 s,72 ℃ 5 min,35 个循环,72 ℃总延伸 10 min,4 ℃停止反应。回收目的片段,分别通过限制性酶切位点 EcoR Ⅰ、Xba Ⅰ 和 Hind Ⅲ 与 pMD19-T 克隆载体连接,构建得到质粒 pMD-sRNA-up 和 pMD-sRNA-down。将测序正确的质粒 pMD-sRNA-up/down 和敲除载体 pK18mobsacB 用限制性内切酶(EcoR Ⅰ 和 Xba Ⅰ)进行双酶切,并连接构建得到重组质粒 pKARD。
将 pKARD 电转入嗜水气单胞菌感受态细胞中,涂布于含有 Kan 的平板上,将生长的单菌落分别划线在编号相同的 Kan 抗性平板和含有 10% 蔗糖的 Kan 抗性平板(Kan+ZT)上。接着挑取在 Kan 平板上生长在 Kan+ZT 平板上不生长或者生长能力较弱的单菌落,即为单交换菌株。将筛选出的单菌落接种在 LB 液体培养基中过夜培养,将菌液梯度稀释后涂布于只含有 10% 蔗糖的平板上(ZT),将 ZT 平板上长出的单菌落分别划线在编号相同的 ZT 平板和 Kan 抗性平板上,挑选在 ZT 平板上生长且在 Kan 抗性平板上不生长的菌株,提取该菌株基因组 DNA,利用引物 sRNA-del-F/R 进行 PCR 并测序验证,得到 sRNA 缺失突变体。
(1) 生长曲线测定:取嗜水气单胞菌野生菌株(WT)和 sRNA 缺失菌株(△sRNA)接种于 LB 培养基中,30 ℃、180 r/min 过夜培养。在无菌条件下,取 0.1 μL 的菌液和 300 μL 的 LB 液体培养基以每组设 5 个平行的标准接种于 96 孔板中,用 Bioscreen 生长曲线测定仪(德国 Bioscreen 公司)设置生长条件为 30 ℃、220 Hz 频率,连续培养 40 h,每隔 1 h 测 1 次OD600值。
(2) 运动能力测定:配制琼脂质量分数分别为 0.5%(集群运动,swarming)和 0.35%(泳动,swimming)的固体 LB 培养基。取野生型和△sRNA 接种于 LB 液体培养基,振荡培养至OD600值为 1。吸取 2 μL 菌液分别加至不同琼脂浓度的固体培养基中心,正置于 37 ℃恒温培养箱培养 48 h 后观察菌落直径,每个样品重复 3 次。
(3) 渗透压稳定性测定:取 WT 和△sRNA 菌株,30 ℃、180 r/min 条件下培养至OD600值为 1。取 0.1 μL 菌液接种于 96 孔板中,分别用 300 μL 含 2% 和 4% NaCl 的 LB 液体培养基进行培养,每组设置 5 个平行,用 Bioscreen 生长曲线测定仪设置生长条件为 30 ℃、220 Hz 频率,连续培养 40 h,每隔 1 h 测定 1 次OD600值,生长条件参照 1.2.5 (1)。
(4) 溶血能力测定:利用灭菌棉签蘸取过夜培养的野生型和△sRNA 突变株,划线至 4% 绵羊红细胞血平板上,倒置于 37 ℃恒温培养箱中培养,48 h 后观察其溶血现象,每个样品重复 3 次。
(5) 蛋白酶活测定:将野生型和△sRNA 菌株在 30 ℃、180 r/min 条件下培养至OD600值为 0.6,离心后将上清液通过 0.22 μm 的 MCE 膜过滤。以 1 mL 偶氮酪蛋白(50 mmol/L 的 Tris-HCl 中终质量浓度为 3 mg/mL,pH 值为 7.5)为底物加入 200 μL 上清液中,37 ℃孵育 30 min 后加入预冷的 10% 三氯乙酸终止反应。离心收集上清后分别加入 96 孔板中并用 1 mol/L 的氢氧化钠中和,使用酶标仪测定 400 nm 处吸光度,确定酶活力。
(6) 生物被膜形成能力测定:将过夜培养的OD600值为 0.1 的野生型和△sRNA 菌液分别加入到 24 孔板中,每孔 600μL,用 LB 液体培养基作为空白对照。将 24 孔板放在 30 ℃恒温培养箱中培养 48 h,吸去菌液,用 PBS 清洗 2 遍去除浮菌,加入 600 μL 甲醇固定 30 min。用ddH2O清洗,然后烘干固定。每孔加入 600 μL 质量分数为 1% 的结晶紫染液,室温染色 10 min 后弃去染液,用 PBS 清洗孔板至澄清状态,最后烘干 30 min,观察生物被膜形成情况。为了进一步测定 2 种菌株的生物被膜产量,向 24 孔板的每孔中加入 600 μL 体积分数为 33% 的冰醋酸溶解结合在生物被膜上的染料,酶标仪测量 595 nm 波长下的吸光度,每组设 3 个重复。
(7) 药物敏感性测定:将野生型和△sRNA 菌株于 LB 液体培养基中过夜培养,用 PBS 缓冲液将菌液稀释成合适浓度的菌悬液并用无菌棉签涂布到 LB 固体培养基上,然后在固体培养基表面放置制作好的 20 种抗生素的药敏纸片。于 30 ℃恒温培养 16 - 24 h,并对其抑菌圈的直径进行测量,每组实验 3 个平行。根据抑菌圈直径大小判断药物的敏感性程度,判断标准参考文献 [15]。
(8) 小鼠半致死剂量的测定:将 20 只驯养 7 d 的体质量为 25 g 的 BALB/c 小鼠平均分成 4 笼,每笼 5 只。将 4 笼小鼠再随机分为 2 组:野生型组(2 笼小鼠)、△sRNA 组(2 笼小鼠)。将野生型和△sRNA 菌株接种于 LB 液体培养基中,30 ℃、180 r/min 培养至相应 CFU 所对应OD600值;5 000 r/min、2 min 收集菌体,用 PBS 分别进行梯度稀释,以备腹腔注射用。小鼠腹腔注射菌液后,定时观察并记录小鼠的死亡情况。
1.3 统计分析
本研究的差异统计学意义检验采用单因素方差分析(ANOVA),∗P<0.05表示差异具有统计学意义,∗∗P<0.01表示差异具有高度统计学意义,用 GraphPad Prism 7 软件作图。
2 结果与分析
2.1 嗜水气单胞菌转录组测序分析
使用 Illumina 测序平台进行嗜水气单胞菌 A.hydrophila ATCC 7966 的转录组测序,共获得原始测序序列(raw reads)17 395 644,对原始测序序列进行过滤,除去带接头的、低质量的 reads 后得到的 Clean reads 为 16 970 198,Clean bases 为2.55x10;Q20 碱基百分比为 97.77%,Q30 碱基百分比为 93.66%,表示碱基识别率的正确度较好。碱基 G 和 C 的数量总和占总碱基数量的百分比为 53.39%。结合各项评估数据显示,测序数据较理想,可靠性较高。
2.2 嗜水气单胞菌中非编码 RNA 分析
在本样品中预测得到 4 个 sRNA,其位置、长度及表达水平如表 3 所示。通过 RNAfold 软件对候选的 sRNA 进行二级结构预测。对筛选得到的 4 个 sRNA 进行进一步分析,其中 sRNA04(序列包含有 sRNA03)基因长度最长,而且表达水平最高,FPKM 值最大,因此对其进行后续研究。
通过序列检索发现,sRNA04 位于 A.hydrophila subsp. hydrophila ATCC 7966 基因组中 AHA_4096 和 AHA_4097 之间的基因间隔区(intergenic region,IGR)。AHA_4096 基因预测编码 CopG 家族转录调控因子(CopG transcriptional regulator),AHA_4097 基因预测编码甲基接收趋化蛋白(methyl-accepting chemotaxis protein)。使用 TargetRNA 2.01 软件对候选 sRNA 的靶基因进行预测,筛选到序列特异性P<0.05的目的性靶基因 77 种。
2.3 sRNA 序列保守性分析
通过 NCBI BLAST 搜索分析 sRNA 的核酸序列,Max target sequences 设为 5 000,发现 sRNA 具有较强的物种特异性,仅存在于气单胞菌属中。通过对嗜水气单胞菌、豚鼠气单胞菌(Aeromonas caviae)、动物气单胞菌(Aeromonas bestiarum)、中间气单胞菌(Aeromonas media)、达卡气单胞菌(Aeromonas dhakensis)和杀鲑气单胞菌(Aeromonas salmonicida)的多序列比对分析发现,sRNA 在 5 种气单胞菌中高度保守,序列一致性均高于 99%。
2.4 嗜水气单胞菌的 sRNA 基因缺失突变体(△sRNA)的构建
为了进一步研究嗜水气单胞菌中 sRNA 的生理功能,本研究构建了 sRNA 基因缺失突变体(△sRNA)。首先构建了基因缺失重组质粒 pKARD。泳道 1 是重组质粒 pKARD 的三酶切结果,得到大小约为 5 200 bp 的载体条带、sRNA 上游基因(660 bp)和下游基因(880 bp)条带;泳道 2、3 是重组质粒 pKARD 的双酶切结果,分别获得载体、sRNA 上游基因和 sRNA 下游基因。实验结果证明重组质粒 pKARD 构建成功。△sRNA 的构建:泳道 1 是用 sRNA-del-F/R 引物以野生型菌株的基因组 DNA 为模板进行 PCR 的产物,大小为 804 bp;泳道 2、3、4 是用 sRNA-del-/R 引物以 sRNA 敲除菌株的基因组 DNA 为模板进行 PCR 的产物,大小为 563 bp。野生型菌株与敲除菌株的产物大小相差 241 bp,与预期值相符,进一步测序比对验证了结果的可靠性。上述结果证实本研究成功构建了敲除菌株△sRNA。
2.5 sRNA 生物特性分析结果
为了检测 sRNA 缺失是否会影响嗜水气单胞菌菌株的生长,本研究测定了野生型菌株与 sRNA 敲除菌株持续 40 h 的生长曲线,野生型菌株与△sRNA 菌株的倍增时间均在 15 h,生长稳定期的OD600值约为 1.85,生长速率无明显差异,表明 sRNA 缺失不直接影响嗜水气单胞菌的生长。
细菌泳动和集群运动是以鞭毛为导向的运动行为,可在半固体环境中检测,同时细菌的运动能力也与病原菌的致病性密切相关。对野生型和△sRNA 菌株进行运动能力检测,sRNA 缺失后在 0.35% 和 0.50% 半固体 LB 平板泳动的直径分别为 25.0 mm 和 21.5 mm,与野生型的运动能力无明显差异。
不同渗透压下野生型菌株和△sRNA 菌株的生长情况,野生型和△sRNA 菌株在高渗环境下的生长曲线大致相同,当 NaCl 质量分数为 2% 时,生长平台期OD600值为 1.45,相比正常盐质量分数(OD600值为 1.85)培养下减少 0.4;当 NaCl 质量分数为 4% 时,生长平台期OD600值为 1.02,相比正常盐质量分数(OD600值为 1.85)培养下减少 0.83。这说明在高渗透压条件下野生型和△sRNA 菌株的生长均受到抑制,且随着渗透压的升高,抑制效果越明显,野生型和△sRNA 菌株受抑制的程度没有明显差异。
2.5.4 对嗜水气单胞菌溶血活性及蛋白酶活性的影响
溶血特性与蛋白酶活力是嗜水气单胞菌重要的致病因子,为了探索 sRNA 是否会对致病性嗜水气单胞菌的溶血活性和蛋白酶活性产生影响,利用含有 4% 绵羊红细胞的血平板对菌株的溶血活性进行测定,并检测蛋白酶活性。结果显示,野生型和△sRNA 菌株均能产生明显的溶血现象,没有显著差异。而野生型的蛋白酶活性显著高于△sRNA 菌株的活性,差异具有统计学意义((P<0.01))。
2.5.5 对生物被膜形成能力的影响
通过测定OD595来评估野生型和△sRNA 菌株的生物被膜形成能力,结果表明,野生型菌株具有较强的生物被膜形成能力,sRNA 菌株生物被膜形成能力显著降低((P<0.01))。
2.5.6 药物敏感性测试结果
药敏测试结果显示,与野生型相比,△sRNA 菌株对青霉素和红霉素的敏感性增强,对链霉素的敏感性减弱,对其他 17 种抗生素的敏感性与野生型一致。其中,野生型菌株对青霉素耐药,但△sRNA 菌株对青霉素转为中度敏感,这提示 sRNA 的功能可能与青霉素敏感性有关。
2.5.7 sRNA 对小鼠的致病性评价
嗜水气单胞菌是一种条件致病菌,为了探究 sRNA 是否直接影响其致病性,分别向小鼠体内注射 3 种不同剂量的野生型和△sRNA 菌株,分别为4.5×107、3.0×107和1.5×107CFUs。感染野生型菌株剂量为3.0×107CFU的小鼠表现出厌食、抑郁、虚弱、眼球和屁股出血等临床症状,24 h 内死亡率为 40%,48 h 内死亡率达到 80%。感染野生型菌株剂量为1.5×107CFUs的小鼠 48 h 内死亡率为 40%,临床症状较轻。感染△sRNA 菌株剂量为1.5×107CFUs的小鼠 48 h 内死亡率为 20%,几乎没有表现出临床症状。统计小鼠的死亡率,△sRNA 菌株对小鼠的LD50值(2.1×107CFUs)是野生型菌株(1.7×107CFUs)的 1.23 倍。以上结果表明 sRNA 基因缺失菌株较野生型的致病性有所降低。
3 讨论与结论
非编码小 RNA(sRNA)在细菌的整个生命活动中起着重要的调控作用,主要是帮助细菌快速感知并适应环境变化、调控细菌代谢、控制群体感应和生物被膜形成以及调控毒力基因表达和细菌致病力。本研究基于嗜水气单胞菌的转录组测序数据,预测得到 4 个非编码 RNA,以其中一段 sRNA 为研究对象,构建得到 sRNA 基因缺失突变体(△sRNA),并对△sRNA 的生理特性和致病性进行分析。
研究显示,敲除 sRNA 基因后,突变体的生物被膜形成能力显著下降,表明 sRNA 的敲除对嗜水气单胞菌生物被膜形成具有显著的抑制作用。细菌生物被膜的形成是一个复杂的生理过程,是细菌适应环境的重要机制。以往研究证实,sRNA 在生物被膜形成过程中起着重要的调控作用。Andreassen 等研究发现,sRNA 通过影响大肠杆菌形成生物被膜的关键转录因子 CsgD 的表达,从而影响其生物被膜的形成;Falcone 等研究发现,铜绿假单胞菌(Pseudomonas aeruginosa)和肠炎沙门菌(Salmonella enteritidis)中有多种 sRNA,可通过调控与生物被膜形成相关的蛋白质影响细菌生物被膜的形成。
革兰氏阴性菌细胞壁外膜上存在外膜蛋白(OMPs),这种蛋白是细菌适应生存环境所必需的,具有良好的免疫原性,如霍乱弧菌(Vibrio cholerae)和福氏志贺氏菌(Shigella flexneri)中某些 sRNA 可以抑制外膜蛋白的合成。此外 sRNA 还会影响细菌体内许多蛋白质的合成,改变细胞必须的营养物质,如细菌代谢所必须的铁离子,它是细胞内许多生化反应重要的辅助因子,细菌中某些 sRNA 对铁代谢过程中多个基因的表达都具有调控作用。如大肠杆菌中的 sRNA - RhyB,它能够调节铁吸收调节蛋白(Fur),RhyB 是细菌感受环境中铁离子浓度下降时表达的一种 sRNA,当 RhyB 表达时,它能够与需铁蛋白的 mRNA 部分序列结合,降低 mRNA 的稳定性和转录效率,减少需铁蛋白合成,从而维持细菌细胞内部的铁稳态。
嗜水气单胞菌 sRNA 基因的缺失降低了菌体的生物被膜形成能力、蛋白酶活性和致病性,但是对菌体的生长能力、运动能力、溶血能力以及对高渗环境的适应能力没有显著影响。宋婷等的研究也发现,短小芽孢杆菌非编码 RNA 基因缺失突变体在生长曲线和胞内蛋白表达方面与野生型菌株无显著差异。因此本研究推断 sRNA 可能通过影响嗜水气单胞菌的生物被膜形成从而影响其致病能力。
针对 sRNA 的功能本研究对 sRNA 基因的处理方式还存在不足。在后续工作中,一方面可以尝试上调 sRNA 基因的表达量,国外有研究表明,在致病性金黄色葡萄球菌中诱导非编码 RNA - sprC 的表达会导致该菌的致病性减弱;另一方面,sRNA 通常是细菌受到胁迫的环境下产生的调控因子,可能在特定的生理功能中影响到细菌表型的变化,这些在本研究中还未涉及。
王嘉璐;蔡彤璇;陈甜梦;李 娜;刘瑶瑶;赵宝华;刘 东,河北师范大学生命科学学院,202501
